

Maquinas moleculares Parte 2
Para Explicar la Vida
Aunque no se han citado aquí muchos detalles de la bioquímica de la visión, hemos pretendido demostrar con este vistazo que, últimamente, esto es lo que significa “explicar” la visión. Éste es el nivel de explicación que la ciencia biológica eventualmente debe perseguir. Para poder decir que se comprende alguna función, se deben dilucidar todos los pasos relevantes en el proceso. Los pasos relevantes en el proceso biológico ocurren, en última instancia, al nivel molecular, de manera que la explicación satisfactoria de un proceso biológico, tales como la vista, la digestión o la inmunidad, deben incluir una explicación molecular. Ahora que la caja negra de la visión ha sido abierta, ya no es suficiente, una “explicación evolucionista” para aclarar “solamente” las estructuras anatómicas de los ojos como un todo, como hizo Darwin en el siglo XIX y como los propagandistas de la evolución hacen hoy.
La anatomía es, sencillamente, irrelevante. También lo es el registro fósil. No importa si el registro fósil es o no consistente con la teoría evolucionista, como no importaba en la física que la teoría de Newton fuese consistente con la experiencia cotidiana. El registro fósil no tiene nada que decirnos sobre, digamos, cómo las interacciones entre el 11-cisretinol con la rodopsina, la transducina y la fosfodiesterasa podrían haberse desarrollado paso a paso. Tampoco los patrones de biogeografía importan, o la genética de poblaciones, o las explicaciones que la teoría de la evolución ha ofrecido para explicar los órganos rudimentarios o la abundancia de especies.
“Cómo es que el nervio viene a ser sensitivo a la luz difícilmente nos importan más que cómo se originó la vida misma”, dijo Darwin el en siglo XIX.
Pero ambos fenómenos han atraído el interés de la bioquímica moderna. La historia de la lenta parálisis de investigación del origen de la vida es muy interesante, pero no tenemos espacio para comentarlo aquí. Es suficiente señalar que actualmente los estudios del origen de la vida se han disuelto en una cacofonía de modelos en conflicto, cada uno poco convincente, seriamente incompleto e incompatible con los demás modelos.
En privado aún la mayoría de los biólogos evolucionistas admitirán que la ciencia no tiene explicación para el origen de la vida.(5) El propósito de este trabajo es el de demostrar que los mismos problemas que acosan la investigación sobre el Origen de la Vida, también lo hacen con los esfuerzos para explicar cómo pudo venir a existir cualquier sistema bioquímico complejo. La Bioquímica ha revelado un mundo molecular que resiste tenazmente ser explicado por el mismo sistema que ha sido aplicado por largo tiempo al nivel de todo el organismo. Ninguna de las cajas negras de Darwin —el origen de la vida o el origen de la visión u otro sistema bioquímico complejo— ha sido demostrado por su teoría.
Complejidad Irreductible
En el Origen de la Especies Darwin dijo:(6)
"Si se pudiera demostrar que cualquier órgano complejo existió que no pudiera haber sido formado por numerosas y sucesivas modificaciones pequeñas, mi teoría se vendría abajo completamente".
Un sistema que llena los requisitos de Darwin es uno que exhibe complejidad irreductible. Por complejidad irreductible quiero significar a un único sistema compuesto de varias partes que interactúan entre sí y que contribuyen a su función básica, y donde la eliminación de cualquiera de estas partes provoca que el sistema deje de funcionar. Un sistema irreductiblemente complejo no puede ser producido gradualmente por modificaciones leves sucesivas de un sistema precursor, ya que cualquier precursor de un sistema irreductiblemente complejo, por definición, no funciona. Ya que la selección natural requiere una función para seleccionar, un sistema biológico irreductiblemente complejo, si es que existe tal cosa, tendría que surgir como una unidad integrada para tener algo sobre lo que actuar. Es casi universalmente admitido que tal súbito evento sería irreconciliable con el gradualismo que Darwin propuso. En este punto, sin embargo, “irreductiblemente complejo” es sólo un término cuyo poder reside principalmente en su definición. Debemos ahora preguntarnos si alguna cosa real es de hecho complejamente irreductible, y, si lo hay, preguntarnos si también existe algún sistema biológico irreductiblemente
complejo.
Considere la ratonera (Figura 3). Las ratoneras que mi familia utiliza en nuestro hogar para lidiar con los indeseables roedores, consisten de una serie de partes. Éstas son: (1) una plataforma plana de madera que sirve como base; (2) un “martillo” de metal que realiza la acción propiamente de aplastar al ratoncito; (3) un resorte de metal con extremos extendidos que ejercen presión sobre la plataforma y el martillo cuando la ratonera está “cargada”; (4) un “gatillo” sensible que se activa cuando se le ejerce una leve presión y (5) una barra de metal que mantiene en su posición al martillo cuando la ratonera está cargada y que conecta con el gatillo. También hay grapas y tornillos que mantienen al sistema en su sitio.

Fig 3.Una ratonera. Si falta cualquiera de sus partes, la ratonera no funciona.
Si uno de los componentes de la ratonera (la base, el martillo, el resorte, el gatillo o la barra de metal) falta, entonces la trampa no funciona. En otras palabras, la sencilla pequeña ratonera no tiene la capacidad de capturar al ratón hasta que todas las partes están en su lugar.
Porque la ratonera está necesariamente compuesta de diferentes partes, es “irreductiblemente compleja”. Por lo tanto, existe un sistema irreductiblemente complejo.
Máquinas Moleculares
Ahora, ¿existe algún sistema bioquímico que sea irreductiblemente complejo? Sí, de hecho existen muchos. Anteriormente hablamos sobre las proteínas. En muchas estructuras biológicas las proteínas son sencillamente componentes de máquinas moleculares aún mayores. Como el tubo, los alambres, tuercas y tornillos en el caso del aparato de televisión, muchas proteínas son partes de estructuras que sólo funcionan cuando básicamente todos los componentes están ensamblados. Un buen ejemplo de esto lo es el cilio.(7)
Los cilios son organelos parecidos a cabellos que existen en la superficie de las células de muchos animales y plantas inferiores. Estos sirven para mover líquido sobre la superficie de la célula o para “remar”, moviendo células individuales a través del líquido. En el hombre, por ejemplo, las células epiteliales que recubren el tracto respiratorio poseen (cada célula) unos 200 cilios que se agitan sincrónicamente para mover la mucosidad hacia la garganta para su eliminación.
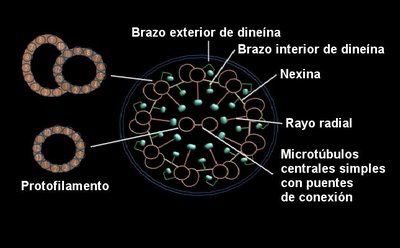
Fig. 4 Vista seccional de un cilio.
Cada cilio está compuesto de un grupo de fibras cubierto por una membrana, que se llama axonema. Un axonema contiene un anillo de 9 microtúbulos dobles rodeando a dos microtúbulos centrales. Cada doblete periférico consiste de un anillo de 13 filamentos (sub-fibra A) fundidos a otra parte compuesta de 10 filamentos (sub-fibra B). Los filamentos de los microtúbulos están compuestos por dos proteínas llamadas alfa y beta - tubulina. Los 11 microtúbulos que forman el axonema se mantienen juntos por tres tipos de conectores: Las sub-fibras A están unidas a los microtúbulos centrales por conexiones radiales [como los de las bicicletas]; los dobletes periféricos adyacentes están unidos por conectores que consisten de una proteína altamente elástica llamada nexina y los microtúbulos centrales están unidos por un puente conector. Finalmente, cada sub-fibra A tiene dos “brazos”, uno exterior y otro interior, ambos conteniendo la proteína “dineína”.

Pero, ¿cómo trabaja el cilio? Los experimentos indican que el movimiento ciliar resulta del “correr” de los brazos de “dineína” sobre un microtúbulo por encima de una sub-fibra B adyacente de un segundo microtúbulo, de manera que los dos microtúbulos se deslizan alternadamente (Figuras 5a y 5b). Todo esto activado por energía química. Sin embargo, los puentes transversales entre los microtúbulos en un cilio intacto previenen que los microtúbulos adyacentes se deslicen uno sobre el otro más allá de una pequeña distancia. Estos puentes transversales, por lo tanto, convierten al movimiento inducido por la dineína en un movimiento de batido de todo el axonema.

Fig. 5b Dibujo esquemático de parte de un cilio. La transmisión de fuerza de la proteína motor, dineína, pegada a un microtúbulo contra la sub-fibra B de un microtúbulo adyacente, causa que las fibras se deslicen uno sobre la otra, adelantándose alternadamente. La proteína unificadora flexible, nexina, convierte el movimiento deslizante en un movimiento de batido.
Ahora, descansemos y repasemos el trabajo del cilio y consideremos lo que esto implica. Los cilios están compuestos de por lo menos 6 proteínas: alfa-tubulina, beta-tubulina, dineína, nexina, conexiones radiales y una proteína central en puente. Éstas se combinan para desarrollar una tarea, el movimiento ciliar, y todas estas proteínas deben estar presentes para que el cilio funcione. Si las tubulinas están ausentes, entonces los filamentos no se deslizan; si la dineína falta, entonces los cilios se quedan rígidos y quietos; si la nexina o alguna otra de las proteínas conectoras faltan, entonces el axonema se deshace cuando los filamentos se deslicen.
Lo que vemos en el cilio, entonces, no sólo es una profunda complejidad, sino que también es una complejidad irreductible a escala molecular. Recuerde que definimos “complejidad irreductible” como un aparato que requiere varios diferentes componentes para que el todo funcione. Mi ratonera debe tener una base, el martillo, un resorte, y una barra sujetadora, todo funcionando coordinadamente, de modo que la ratonera lleve a cabo su trabajo. De manera similar, el cilio, tal y como está constituido, debe tener los filamentos deslizantes, las proteínas conectoras y las proteínas motoras para que funcione. En la ausencia de alguno de estos componentes, el aparato es inservible.
Los componentes del cilio son moléculas específicas. Esto quiere decir que no se pueden invocar aquí “cajas negras”; la complejidad del cilio es final, fundamental. Y, de la misma manera que los científicos (cuando comenzaron a descubrir las complejidades de la célula) se dieron cuenta de lo tonto que es pensar que la vida surgió espontáneamente en un sólo paso o a través de varios pasos desde el fango del océano, de la misma manera nos damos cuenta que el complejo cilio no puede conseguirse en un sólo paso o en una serie de pasos. Pero como la complejidad del cilio es irreductible, entonces no puede tener precursores funcionales. Ya que el cilio irreductiblemente complejo no puede tener precursores funcionales no puede haber sido producido por la selección natural, que requiere de un funcionamiento contínuo. La selección natural es impotente cuando no hay función que seleccionar. Podemos ir más allá y decir que, si el cilio no puede producirse por selección natural, entonces el cilio fue diseñado.
Click aquí, para pasar a:_Maquinas moleculares Parte 3